(2/8)
2.タンパク質立体構造の予測問題それでは、本題に入らせていただきます。
タンパク質立体構造の予測問題ということで、本日は、以下の順序で話させていただきます。
- 「なぜ構造予測問題が難しいのか」
その理由についてお話しします。- 構造予測の研究史
研究の歴史もずいぶんと長いので、その経過をかいつまんでお話しします。- アブイニシオ予測の可能性
構造予測の研究方法の中で、特に注目されているアブイニシオ予測の可能性についてお話しします。- 予測問題が解決すれば・・!
最後に、タンパク質立体構造の予測問題が解決するとどうなるのか、その影響についてお話しします。
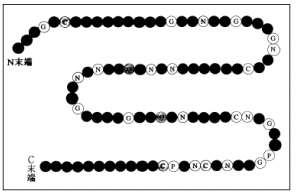
2-1.なぜ構造予測問題は難しいのか?皆さんもよくご存知のタンパク質は、DNAからアミノ酸の配列として翻訳すると、画面5に示す通り「ひも状分子」(鎖状高分子)と呼ばれる1次元の高分子であり、N末端から始まってC末端で終わるという方向性を持っています。そして、アミノ酸の配列の順番が違うと、出来上がる立体構造も異なるという大きな特徴を持っています。
画面6は、今から40年ほど前の1960年に、X線結晶解析で初めて解かれたミオグロビンの立体構造です。当時の人は、ミオグロビンはもう少しカチっとした結晶のようなものだと予想していたのに対し、結果はご覧の通り一見不規則に見えるものでしたから、非常に驚きました。そして、このことから、なぜアミノ酸の配列だけで別の構造になってしまうのかということを意識しだした人が非常に増えたわけです。そこから、現在に至る「予測問題」〜コンピュータにアミノ酸の配列だけを与えて、計算の上で立体構造を予測する〜 が意識されだしました。
画面5
画面6
画面7
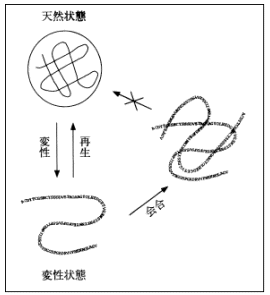
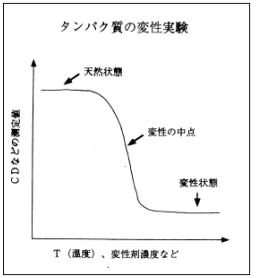
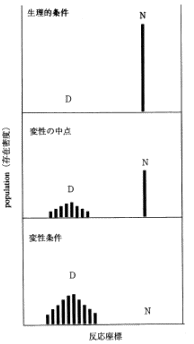
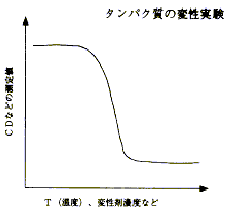
画面7をご覧下さい。これは、温度を上げると天然状態のものが壊れてランダムコイルに戻り(変性)、逆に温度を丁寧に下げてやるともとの天然状態に戻る (再生) という関係を表した図です。この図からも分かるように、タンパク質は、このような可逆的な過程で「安定構造」であることが解明されています。ならば、コンピュータで安定構造を分子力学的にエネルギー計算すれば、立体構造は求まるはずだということは当初から分かっていました。しかし実際に解こうとすると非常に難しく、「予測問題」というものが論文などで議論されだしてから30年以上経つ今でも、未だに完全な解決策はでていません。その理由は、タンパク質の特性に大きく関係しています。画面8は、タンパク質の温度を上げていった際の変性曲線ですが、温度を上げると、天然状態からトランディションして変性状態に戻ります。別の見方をすると、画面9から分かるように、環境状態を生理的な条件にすれば、ポピュレーション (population:存在密度) は天然の状態(N)にあるが、逆に変性しやすいような環境状態にすれば、ポピュレーションは全て変性(D)に偏るということです。
画面8
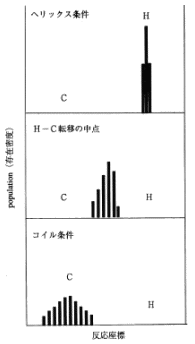
途中過程の "変性の中点" といわれているところでは、実際に何が起こっているかを見てみると、ご覧の通り、ポピュレーションが天然状態と変性状態の2つに分かれています。重要なのは、NもDも両方あり、両者の中間状態には留まらないという点です。このように、半分が壊れて半分が出来ているというような、そのような例は他の多数の分子をみても見当たりません。NかDかどちらか一方である例が多数なのです。タンパク質のフォールディング (folding:折りたたみ過程) におけるこのような特徴は、「協同性 (cooperativity) 」と呼ばれています。
画面9
画面10
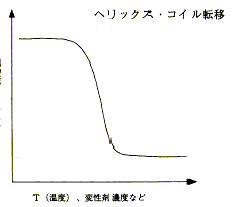
少し話はそれますが、それでは、変性の中点ではNかDかどちらか一方であることが当たり前かというと、違う例もたくさんあります。例えばDNAのヘリックス・コイル転移は、天然状態がヘリックス(二重螺旋)で、変性の結果コイル状になるのですが、この場合の中点は、画面10の通りポピュレーションが中間状態にあり、先ほどのタンパク質とは全く異なります。ある部分ではヘリックスが残っていて、ある部分ではほどけていて、それが入り混じっているという状態です。このように、タンパク質には「協同性」という特性があるわけですが、画面11と12を見比べるとお分かりの通り、面白いことに、変性曲線では、タンパク質もヘリックス・コイル転移もどちらも同じように見えます。しかし、中身は全く違うということがお分かりいただけたと思います。
画面11
画面12